Lecture 29. Mendelian discovery of genes
Mendelian inheritance (or Mendelian genetics or Mendelism) is a set of primary tenets relating to the transmission of hereditary characteristics from parent organisms to their children; it underlies much of genetics. They were initially derived from the work of Gregor Mendel published in 1865 and 1866 which was "re-discovered" in 1900, and were initially very controversial. When they were integrated with the chromosome theory of inheritance by Thomas Hunt Morgan in 1915, they became the core of classical genetics.
The laws of inheritance were derived by Gregor Mendel, a 19th century [1] monk conducting hybridization experiments in garden peas (Pisum sativum). Between 1856 and 1863, he cultivated and tested some 28,000 pea plants. From these experiments he deduced two generalizations which later became known as Mendel's Laws of Heredity or Mendelian inheritance. He described these laws in a two part paper, "Experiments on Plant Hybridization" that he read to the Natural History Society of Brno on February 8 and March 8, 1865, and which was published in 1866.[2]
The principles of heredity were written by the Augustinian monk Gregor Mendel in 1865. Mendel discovered that by crossing white flower and purple flower plants, the result was a hybrid offspring. Rather than being a mix of the two, the offspring was purple flowered. He then conceived the idea of heredity units, which he called "factors", one of which is a recessive characteristic and the other dominant. Mendel said that factors, later called genes, normally occur in pairs in ordinary body cells, yet segregate during the formation of sex cells. Each member of the pair becomes part of the separate sex cell. The dominant gene, such as the purple flower in Mendel's plants, will hide the recessive gene, the white flower. After Mendel self-fertilized the F1 generation and obtained the 3:1 ratio, he correctly theorized that genes can be paired in three different ways for each trait; AA, aa, and Aa. The capital A represents the dominant factor and lowercase a represent the recessive.
Mendel stated that each individual has two factors for each trait, one from each parent. The two factors may or may not contain the same information. If the two factors are identical, the individual is called homozygous for the trait. If the two factors have different information, the individual is called heterozygous. The alternative forms of a factor are called alleles. The genotype of an individual is made up of the many alleles it possesses. An individual's physical appearance, or phenotype, is determined by its alleles as well as by its environment. An individual possesses two alleles for each trait; one allele is given by the female parent and the other by the male parent. They are passed on when an individual matures and produces gametes, egg and sperm. When gametes from the paired alleles separate randomly, each gamete receives a copy of one of the two alleles. The presence of an allele doesn't promise that the trait will be expressed in the individual that possesses it. In heterozygous individuals, the only allele that is expressed is the dominant. The recessive allele is present but its expression is hidden.
Mendel summarized his findings in two laws; the Law of Segregation and the Law of Independent Assortment.
Now when we know the mechanisms of meiosis, one can conclude that the two abovementioned Mendelian laws are direct consequences of the assortment laws of chromosomes in meiotic cell division, and the Mendelian “factors” are today’s genes.
A good description of Mendel’s pea crosses and his detail experiments was presented in MITOPENCOURSEWARE (PDF).
Lecture 30. Gene linkage
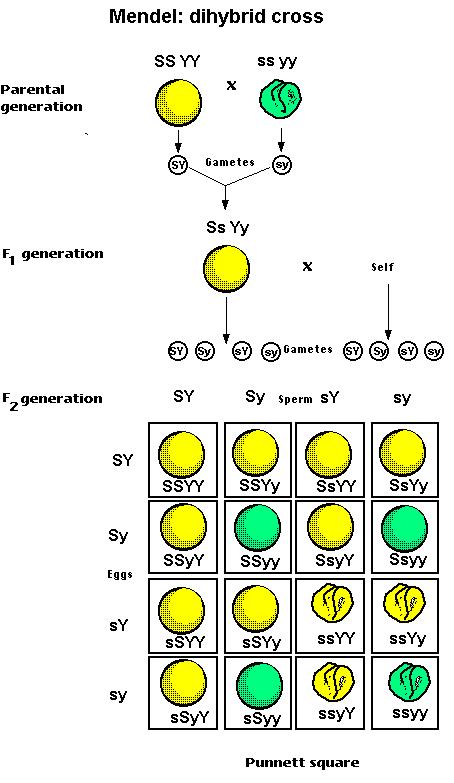
Gregor Mendel analyzed the pattern of inheritance of seven pairs of contrasting traits in the domestic pea plant. He did this by cross-breeding dihybrids; that is, plants that were heterozygous for the alleles controlling two different traits.
Mendel then crossed these dihybrids. If it is inevitable that round seeds must always be yellow and wrinkled seeds must be green, then he would have expected that this would produce a typical monohybrid cross: 75% round-yellow; 25% wrinkled-green. But, in fact, his mating generated seeds that showed all possible combinations of the color and texture traits.
- 9/16 of the offspring were round-yellow
- 3/16 were round-green
- 3/16 were wrinkled-yellow, and
- 1/16 were wrinkled-green
Finding in every case that each of his seven traits was inherited independently of the others, he formed his "second rule", the Rule of Independent Assortment:
The inheritance of one pair of factors (genes) is independent of the inheritance of the other pair. Today we know that this rule holds only if the genes are on separate chromosomes.
Mendel was lucky in that every pair of genes he studied met one requirement or the other. The table shows the chromosome assignments of the seven pairs of alleles that Mendel studied. All of these genes showed independent assortment, and they were inherited on separate chromosomes. With the rebirth of genetics in the 20th century, it quickly became apparent that Mendel's second rule does not apply to many matings of dihybrids. In many cases, two alleles inherited from one parent show a strong tendency to stay together as do those from the other parent. This phenomenon is called linkage.
So, gene linkage is the physical relationship of genes. Specifically,
linkage means that the genes are on the same chromosome and therefore do not assort independently into gametes --in humans, ovum and spermatozoa-- during meiosis.
Because of this co-transmittance, the traits associated with the genes do not segregate between two daughter cells, following crosses between the parental cells, as predicted by Mendelian genetics.
The genes of most organisms can exist in different forms, called alleles, in a population. If the organism has identical alleles of a gene on each of its homologous chromosomes, it is called homozygous. If the alleles are different, it is called heterozygous. During the cell division process, a
separation of nuclear material into gametes occurs via meiosis. If an organism is heterozygous, two kinds of gametes are produced; if homozygous, it produces only one kind of gamete. At fertilization the male and female gametes combine and the random process that creates different units put the gametes into various combinations. The ratio of the appearance of the observed traits, or phenotypes, produced by the pattern of separation of the dominant and recessive genes for that trait was predicted by Gregor Mendel following painstaking work and observation of the crosses between pea plants.
However, early in the twentieth century, William Bateson and Reginald Crundall Punnett, two British geneticists, observed that sometimes the expected Mendelian ratio of phenotypes did not occur. Their best explanation was that in some manner the phenotypic classes, the alleles, were coupled, and so did not sort independently into gametes. Proof of their explanation was provided by Thomas Hunt Morgan, using Drosophila eye color as the examined trait.
Morgan observed that test crosses between mutants in eye color and wing development deviated from the expected Mendelian 1:1:1:1 ratio for independent assortment. The observed ratio was, rather, consistent with the non-independent segregation of two genes that were close to each other on the same chromosome.
Linked genes do not observe the genotypic or phenotypic relationships predicted by Mendelian crosses that assume independent assortment of chromosomes and genes. In a cross the parental generation is designated P1 and the first generation of offspring are designated F1(first filial generation), and the offspring resulting from the fertilization between individuals of the F1 generation are called the F2 (second filial generation). When the F1 and F2 ratios deviate from the predicted Mendelian ratios, this is evidence of gene linkage.
The linkage of genes is used to generate so-called
linkage maps which give a measure of the distance between genes on a chromosome. The
linkage map technique, which is based on the use of the percentage of recombinants, in which crossing over of DNA and expression of traits due to gene
linkage has occurred, was devised in 1911 by Alfred Henry Sturtevant, an undergraduate student of Morgan's. The technique remains in use today as a means of producing an index of the distance between two genes.
To see concrete experiments for analyzing as well as for applying gene linkage, click over to MITOPENCOURSEWARE (PDF) for the complementaton test and gene function; click to (PDF) for tests of gene position, starting with the position of genes on chromosomes in general; click to (PDF) for experiments to map genes relative to one another on sex chromosomes, and, finally, click to (PDF) for mapping genes on autosomes by test-cross and other measures.
Lecture 31. Genetic complex traits
Genetic complex traits refer to those traits which are determined by either many genes or vice versa a single gene influences multiple phenotypictraits.
This lecture note will present the main types of the traits of the kind-- Pleiotropy, Polygenic Inheritance, Genetic Heterogeneity, Twinnings and Siblings.
Pleiotropy Pleiotropy occurs when a single gene influences multiple phenotypictraits. Consequently, a new mutation in the gene will have an effect on all traits simultaneously. This can become a problem when selection on one trait favors one specific mutant, while the selection on the other trait favors another mutant. The underlying pleiotropic mechanism is that the gene codes for a product that is, for example, used by various cells, or has a signaling function on various targets.
A classic example of pleiotropy is the human disease PKU (phenylketonuria). This disease can cause mental retardation and reduced hair and skinpigmentation, and can be caused by any of a large number of mutations in a single gene that codes for an enzyme (phenylalanine hydroxylase) that converts the amino acidphenylalanine to tyrosine, another amino acid. Depending on the mutation involved, this results in reduced or zero conversion of phenylalanine to tyrosine, and phenylalanine concentrations increase to toxic levels, causing damage at several locations in the body. PKU is totally benign if a diet free from phenylalanine is maintained.
Antagonistic pleiotropy refers to the expression of a gene resulting in multiple competing effects, some beneficial but others detrimental to the organism.
This is central to a theory of aging first developed by G. C. Williams in 1957.[1] Williams suggested that some genes responsible for increased fitness in the younger, fertile organism contribute to decreased fitness later in life. One such example in male humans is the gene for the hormone testosterone. In youth, testosterone has positive effects including reproductive fitness but, later in life, there are negative effects such as increased susceptibility to prostate cancer. Another example is the p53 gene which suppresses cancer, but also suppresses stem cells which replenish worn-out tissue[2].
Whether or not pleiotropy is antagonistic may depend upon the environment. For instance, a bacterial gene that enhances glucose utilization efficiency at the expense of the ability to use other energy sources (such as lactose) has positive effects when there is plenty of glucose, but it can be lethal if lactose is the only available food source.
Polygenic inheritance Polygenic inheritance is a pattern responsible for many features that seem simple on the surface. Many traits such as height, shape, weight, color, and metabolic rate are governed by the cumulative effects of many genes. Polygenic traits are not expressed as absolute or discrete characters, as was the case with Mendel's pea plant traits. Instead, polygenic traits are recognizable by their expression as a gradation of small differences (a continuous variation). The results form a bell shaped curve, with a mean value and extremes in either direction.
Height in humans is a polygenic trait, as is color in wheat kernels. Height in humans is NOT discontinuous. If you line up the entire class, a continuum of variation is evident, with an average height and extremes in variation [very short (vertically challenged) and very tall (vertically enhanced)]. Traits showing continuous variation are usually controlled by the additive effects of two or more separate gene pairs. This is an example of polygenic inheritance. The inheritance of EACH gene follows Mendelian rules.
- 1. Traits are usually quantified by measurement rather than counting.
- 2. Two or more gene pairs contribute to the phenotype.
- 3. Phenotypic expression of polygenic traits varies over a wide range.
- 1. Height
- 2. SLE (Lupus). (Click here for an article about lupus and genetics.)
- 3. Weight. (Click here for an article about obesity and genetics.)
- 4. Eye Color. (Click here for an article about eye color.)
- 5. Intelligence.
- 6. Skin Color.
- 7. Many forms of behavior.
Click here to see Genetic Heterogeneity, Twinnings and Sibblings described by MITOPENCOURSEWARE.
Lecture 32. Probability and pedigrees
In Mendel’s time he used statistics to account for his observations on his experiments on peas, and, thanks to the results he abtained, he could formulate his two famous laws of genetics-- the Law of segregation and the Law of independent assortment, which were based on statistical segregation ratio 3:1, 9:3:3:1, 1:1:1:1 etc…
Nowadays in genetic research and especially in medical genetic counseling, statistics is needed for calculating the risks of genetic diseases in human pedigrees. The risks in these cases are expressed in terms of so-called probability.
The probability of an event is the chance that it will happen. The probability of tossing a coin to land heads up is roughly ½.
- The probability of an impossible event is 0.
- the probability of a certain event is 1.
- If the probability of event x is p then the probability of 'not x' is 1-p.
- The probability of two independent events ocurring at the same time is the product of their two indivdual probabilities.
- probability of a wrinkled seed is ¼; the probability of a green seed is also ¼, and the probability of being both green and wrinkled is therefore ¼ x ¼ = 1/16.
- The probability of being not wrinkled (i.e. smooth) is 1-¼ = ¾. The probability of being both smooth and green is therefore ¾ x ¼ = 3/16 and so on.
- In the example below about the coefficient of inbreeding of children from first cousin marriages, we considered a number of probabilities of ½ which we multiplied together to reach a final probability of 1/16 that any gene was homozygous by descent.
Autosomal recessive A recessive trait will only manifest itself when homozygous. If it is a severe condition, it will be unlikely that homozygotes will live to reproduce, and thus most occurrences of the condition will be in matings between two heterozygotes (or carriers). An autosomal recessive condition may be transmitted through a long line of carriers before, by ill chance, two carriers mate. Then there will be a ¼ chance that any child will be affected. The pedigree will therefore often only have one 'sibship' with affected members.

Considering any child of a first cousin mating, we can trace through the pedigree the chance that the other allele is the same by common descent. Let us consider any child of generation IV, any gene which came from the father, III3 had a half chance of having come from grandmother II2, a further half chance of being also present in her sister, grandmother II4 a further half a chance of having been passed to mother III4 and finally a half chance of being transmitted into the same child we started from. A total risk of ½ x ½ x ½ x ½ = 1/16.
This figure, which can be thought of as either
- the chance that both maternal and paternal alleles at one locus are identical by descent or
- the proportion of all the individual's genes that are homozygous because of identity by common descent,
is known as the coefficient of inbreeding and is usually given the symbol F.
To see a compact and also clear description of using statiistics for pedigree genetic analysis, click MITOPENCOURSEWARE (PDF).
Lecture 33. Population genetics
If genetics is a science studying structure, function and movement rules of genes, population genetics is the third part of it: a science studying movement rules of gene carriers – chromosomes and their effects and consequences. At least from the genetic point of view, population is a unit of evolution. In terms of breeding practice, populations are plant varieties and animal breeds.
D. S. Falconer (The quote is from Introduction to Quantitative Genetics by D. S. Falconer, 1960, Ronald Press.) wrote:
"A population in the genetic sense, is not just a group of individuals, but a breeding group; and the genetics of a population is concerned not only with the genetic constitution of the individuals but also with the transmission of the genes from one generation to the next. In the transmission the genotypes of the parents are broken down and a new set of genotypes is constituted in the progeny, from the genes transmitted in the gametes. The genes carried by the population thus have continuity from generation to generation, but the genotypes in which they appear do not. The genetic constitution of a population, referring to the genes it carries, is described by the array of gene frequencies, that is by specification of the alleles present at every locus and the numbers or proportions of the different alleles at each locus." (page 6).
In fact population genetics is studying the allele frequency distribution and change under the influence of the four evolutionary forces: natural selection, genetic drift, mutation and gene flow. It also takes account of population subdivision and population structure in space. As such, it attempts to explain such phenomena as adaptation and speciation. Population genetics was a vital ingredient in the modern evolutionary synthesis whose primary founders were Sewall Wright, J. B. S. Haldane and R. A. Fisher, they also laid the foundations for the related discipline of quantitative genetics.
For humans the applications of Mendelian genetics, chromosomal abnormalities, and multifactorial inheritance to medical practice are quite evident. Physicians work mostly with patients and families. However, as important as the work of physicians may be, genes also affect populations, and in the long run their effects in populations have a far more important impact on medicine than the relatively few families each physician may serve. It is important that certain polymorphisms are maintained so that the species may survive, even at the expense of individuals. Genetic polymorphisms often are detrimental to the homozygote, but they allow others of the species to survive. Before medical intervention was possible, populations that lacked the sickle cell anemia allele could not survive in the malaria regions of West Africa. Those that had the sickle cell anemia allele survived, and the gene remains in the population at high frequency today, even though the homozygous recessive phenotype was at a severe disadvantage in the past. The high rate of thalassemia in people of Mediterranean origin, the high rate of sickle cell anemia in people of West African descent, the high rate of cystic fibrosis in people from Western Europe, and the high rate of Tay-Sachs disease in ethnic groups from Eastern Europe may all owe their origin to environmental factors that cause changes in gene frequencies in large populations by giving some advantage to heterozygotes who carry a deleterious allele. Although one may never use the calculations of population genetics in medical practice, the underlying principles should be understood.
To have general understanding of population genetics, click (PDF) for studying Hardy-Weinberg Equilibrium; click (PDF) to see the role of Mutation and Selection in population structure, and click (PDF) for consideration of Inbreeding as a factor influencing the composition of populations.

